Химическая энциклопедия
"ТРАНСЛЯЦИЯ"
Главная - Химическая энциклопедия - буква Т - ТРАНСЛЯЦИЯ
ТРАНСЛЯЦИЯ
(от лат.
translatio-передача), программируемый генами процесс синтеза белка. Посредством
Т. осуществляется реализация генетич. информации нуклеиновых к-т (см. Генетический код
). По совр. представлениям,
исходный ген
в виде ДНК непосредственно транслироваться не может; для
Т. он должен быть сперва транскрибирован-переписан (см. Транскрипция
)-в
форме молекул матричных рибонуклеиновых кислот
(мРНК) и именно последняя
связывается с рибосомой и транслируется. В ходе Т. последовательность
нуклеотидов мРНК определяет последовательность остатков аминокислот в синтезируемом
полипептиде; происходит как бы перевод с языка нуклеотидной последовательности
гена на язык аминокислотной последовательности белка. Считается, что аминокислотная
последовательность синтезируемого на рибосоме полипептида однозначно задает
пространственную структуру белка через процесс сворачивания (фолдинга), идущего
одновременно с Т. (котрансляционное сворачивание) или по ее завершении (посттрансляционное
сворачивание). Кроме того, в формировании конечной структуры функцион. белка
могут принимать участие процессы ко- и посттрансляционных модификаций (см. Модификация белков
). Т. каждой молекулы мРНК
рибосомой разделяется на три четкие последоват. стадии-инициацию, элонгацию
полипептида (собственно
Т.) и терминацию (см. рис. в ст. Рибосома
). Цепь мРНК транслируется
(считывается) по направлению от 5-конца к 3-концу (см. Нуклеиновые кислоты
).
Полипептид элонгируется (растет) от N-конца к С-концу. Т. начинается со строго
фиксир. точки в цепи мРНК (а отнюдь не с начала самой цепи РНК). Точный выбор
рибосомой стартовой точки определяет не только первую (N-концевую) аминокислоту
в синтезируемом полипептиде, но и всю последующую разбивку нуклеотидной последовательности
мРНК на триплеты (кодоны), т.е. правильную фазу (рамку) считывания, и, соотв.,
правильный набор остатков аминокислот в синтезируемом полипептиде. Для этого
существует спец. механизм инициации Т., в к-ром принимают участие: инициаторный
кодон и предшествующая ему последовательность мРНК, малая субчастица рибосомы,
инициаторная метионил-тРНК [N-формилметионил-тРНКр у прокариот (бактерии и синезеленые
водоросли) и метионил-тРНКр у эукариот (все остальные организмы); тРНК-сокр.
обозначение транспортных рибонуклеиновых кислот
], набор белковых
факторов инициации (IF-1, IF-2 и IF-3 у прокариот и около десятка факторов-от
eIF-1 до eIF-4F и eIF-5-y эукариот) и гуанозинтрифосфат (ГТФ), а также АТФ у
эукариот. Перед инициацией рибосома
должна диссоциировать на составляющие ее субчастицы-малую (30S у прокариот и
40S у эукариот) и большую (50S у прокариот и 60S у эукариот): 70S : 30S;
+ 50S 80S : 40S;+60S Своб. малая субчастица
связывает на себе часть факторов инициации (кстати, способствующих вышеуказанной
диссоциации). При участии фактора инициации IF-2 или eIF-2 и ГТФ в комплекс
с малой субчастицей входит инициаторная (формил)метионил-тРНКр (F в нижнем индексе
обозначает, что данная тРНК является инициаторной). У прокариот малая субчастица
рибосомы имеет сродство к короткой полипуриновой (см. Пуриновые основания
)
последовательности мРНК (напр., GAGG; G и А-соотв. остатки гуанозина и аденозина),
находящейся за неск. нуклеотидов перед инициаторным кодовом AUG (реже GUG; U-остаток
уридина), так что рибосомная 30S частица фиксирует эту предынициаторную последовательность
(наз. также последовательностью Шайна-Дальгарно), а антико-дон инициаторной
тРНК взаимод. с инициаторным кодо-ном. У эукариот рибосомная 40S
частица, несущая ряд факторов инициации и метионил-тРНКр, связывается преимущественно
с 5-концом цепи мРНК (как правило, кэпирован-ным), а затем скользит по цепи
в направлении к 3-концу без Т., потребляя АТФ, пока не натолкнется на триплет
AUG, спаривающийся с антикодоном тРНКр и служащий инициаторным кодовом. Т. обр., в обоих случаях
устанавливается стартовая точка Т. (точка отсчета триплетов). Далее фиксированная
на ини-циаторном кодоне малая рибосомная субчастица присоединяет к себе большую
рибосомную субчастицу; это событие сопровождается гидролизом ГТФ на факторе
инициации IF-2 или eIF-2, уходом этого фактора и гуанозиндифосфата (ГДФ) с рибосомной
частицы. Теперь на ишщиаторном кодоне находится полная (70S или 80S) рибосома,
готовая воспринять аминоацил-тРНК (Аа-тРНК), соответствующую следующим нуклеотидным
триплетам мРНК (рис. 1). Элонгационный цикл начинается
с поступления в рибосому Аа-тРНК при наличии там инициаторной (формил)ме-тионил-тРНКр
(сразу после вышеописанной инициации) или пептидил-тРНК (если рассматривать
любой промежут. шаг стадии элонгации) (рис. 2). Предварительно вне рибосомы
Аа-тРНК взаимод. со спец. белком, наз. фактором элонгации Tu (EF-Tu)
у прокариот или eEF-1 у эукариот; для взаимод. необходимо участие в этом комплексе
молекулы ГТФ: Аа-тРНК + EF-Tu(eEF-l) + ГТФ :!
Аа-тРНК · EF-Tu(eEF-l)·ГТФ 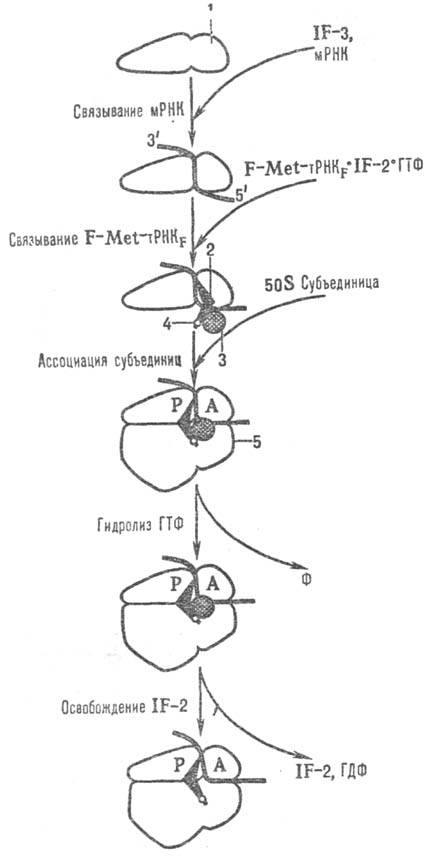
Рис. 1. Схема последовательности
событий в процессе инициации трансляции у прокариот: 1-малая субъединица рибосомы;
2-инициаторная тРНК; 3-фактор инициации IF-2; 4-N-формилметионил (сокр. F-Met);
5-большая субъединица рибосомы; P и А-тРНК-связывающие участки рибосомы,
F-фосфат. 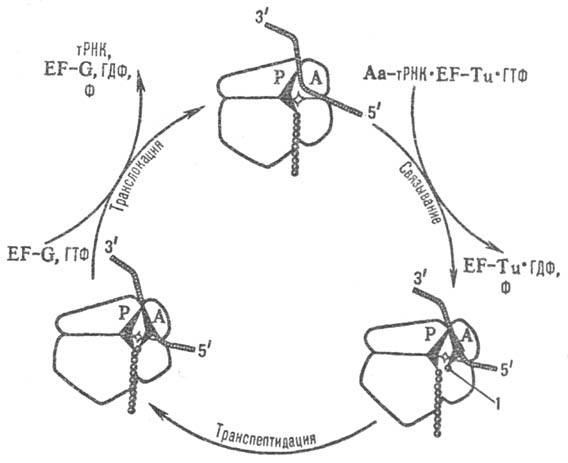
Рис. 2. Схема элонгационного
цикла рибосомы; EF-Tu и EF-G-факторы элонгации, Аа-тРНК-аминоацил-тРНК,
1 -остаток аминокислоты, связанной с тРНК, F-фосфат. 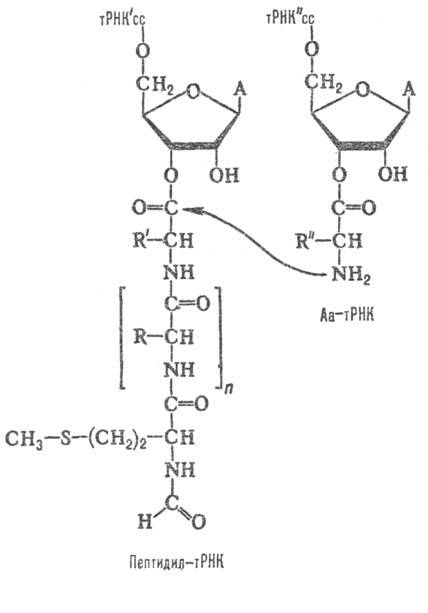
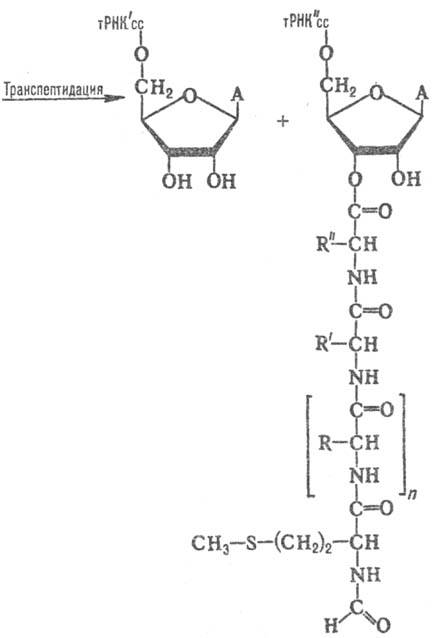
Рис. 3. Р-ция транспептидации,
катализи-руемая рибосомой; cc-два концевых остатка цитидина в тРНК, А-остаток
аде-нина. На самом деле, ввиду избытка
белка ЕР-Тu или eEF-1 в клетке, все Аа-тРНК, за исключением инициаторной,
присутствуют в цитоплазме в виде таких тройственных нуклеопротеидных комплексов.
Комплекс поступает в рибосому и связывается с ее т. наз. А-участком, если
там присутствует вакантный кодон, комплементарный антико-дону Аа-тРНК. При этом
др. тРНК-связывающий участок рибосомы (Р-участок) занят либо инициаторной (фор-мил)метионил-тРНКр
(в первом элонгационном цикле), либо пептидил-тРНК (во всех последующих элонгационных
циклах). Связывание тройственного
комплекса с А-участком рибосомы индуцирует его ГТФ-азную активность, и
ГТФ, входящий в комплекс, гидролизуется: ГТФ + Н2О :
ГДФ + Н3РО4 В результате этого EF-Tu
(eEF-1) теряет высокое сродство к рибосоме и к Аа-тРНК и вместе с ГДФ покидает
рибосому. Теперь Аа-тРНК оказывается
в А-участке рибосомы без своего белкового партнера, бок о бок с присутствующей
в Р-участке инициаторной (формил)метионил-тРНКр (или пептидил-тРНК). Аминокислотный
остаток Аа-тРНК и сложноэфирная группа (формил)метионил-тРНКр (или пеп-тидил-тРНК)
локализуется в пептидил-трансферазном центре рибосомы. Аминогруппа аминокислотного
остатка атакует карбонил сложноэфирной группы, в результате чего происходит
р-ция транспептидации: (формил)метионильный (или пептидильный) остаток переносится
на аминогруппу Аа-тРНК, с образованием пептидной связи, а инициаторная тРНКF
(или тРНК, с к-рой был связан пептид) оказывается деацилированной (рис. 3). Непосредственно после р-ции
транспептидации деацили-рованная тРНК занимает Р-участок рибосомы, а новообразованная
пептидил-тРНК-А-участок (см. рис. 2). Завершающая фаза цикла наз. транслокацией.
Она катализируется крупным мономерным белком, обозначаемым как фактор элонгации
G (EF-G) у прокариот или eEF-2 у эукариот, с использованием молекулы ГТФ. Фактор EF-G(eEF-2), ассоциированный
с ГТФ, взаимод. с претранслокационной рибосомой (пептидил-тРНК в А-участке,
деацшшрованная тРНК в Р-участке), и в образующемся комплексе происходит удаление
деацилирован-ной тРНК из Р-участка, переход пептидил-тРНК из А-участ-ка в Р-участок
и передвижение вместе с ней кодона мРНК из А-участка в Р-участок; в вакантном
А-участке устанавливается очередной кодон мРНК. В образовавшемся посттранс-локационном
комплексе индуцируется ГТФ-азная активность, ГТФ гидролизуется до ГДФ и Н3РО4,
в результате чего EF-G(eEF-2) с ГДФ теряет сродство к рибосоме и освобождается.
Цикл завершен. Теперь рибосома способна
воспринять на свой вакантный А-участок с очередным кодовом новую, соответствующую
этому кодону Аа-тРНК в комплексе с EF-Tu (eEF-1) и ГТФ. Осуществляется
следующий цикл элонгации. Трансляция мРНК состоит в повторении таких циклов,
в каждом из к-рых прочитывается один триплет (кодон) мРНК, синтезируется одна
пептидная связь и гидролизуются две молекулы ГТФ. Элонгация полипептида продолжается
до тех пор, пока рибосома не натолкнется на один из нуклеотидных триплетов,
не кодирующих аминокислоту, т.е. не комплементарных антикодону какой-либо тРНК.
Это триплеты UAA, UAG и UGA, наз. терминаторными или стопкодонами. Тогда включается
механизм терминации: терминаторный триплет, оказавшийся в А-участке рибосомы,
узнается не молекулой тРНК, а спец. белком (фактором терминации RF) с участием
ГТФ; фактор терминации, связавшийся с рибосомой, индуцирует гидролиз сложноэфирной
связи между полипептидом и тРНК в молекуле пептидил-тРНК, расположенной в Р-участке
рибосомы. Гидролиз катализируется
пептидил-трансферазным центром рибосомы, но в данном случае сложноэфирный карбо-нил
атакуется не аминогруппой Аа-тРНК, как при транспептидации, а молекулой воды.
Готовый полипептид освобождается из рибосомы, а рибосома подвергается действию
дополнит. факторов, лишается последней деацилированной тРНК, диссоциирует (при
участии факторов инициации) на субчастицы и покидает мРНК. Своб. рибосомные
субчастицы опять готовы инициировать Т. новой цепи мРНК или нового цистрона
(кодируемого участка) на цепи той же полицистронной мРНК. Реализация генетич. информации
через Т. подвержена ошибкам. Ошибки в Т. могут быть двоякого рода. Ошибки первого
рода-это т. наз. ложное считывание кодона, когда очередная Аа-тРНК при поступлении
в А-участок рибосомы связывается не соответствующим ей коданом. Определенная
вероятность такого захвата (оцениваемая величинами порядка 10-3-10-4)
обычно характерна для тРНК, несущих антикодон, частично комплементарный кодону.
Результатом является замена правильного аминокислотного остатка на неправильный
в соответствующем положении полипептидной цепи. Вариантом этого рода ошибок
является связывание, напр. триптофанил-тРНК, с терминаторным кодо-ном в А-участке
и, как следствие,-продолжение элонгации за пределы терминаторного кодона, т.
е. синтез удлиненного полипептида. Уровень ошибок зависит как от ряда внеш.
факторов (напр., возрастает при голодании или в присут. этанола), так и от структуры
самой рибосомы. В частности, известны мутации рибосомных белков, увеличивающие
или уменьшающие, избирательность рибосомы при поступлении Аа-тРНК в А-участок
и соотв. уменьшающие или увеличивающие уровень опшбок этого рода. Ошибки др. рода-это т.
наз. сдвиг рамки считывания мРНК, когда при транслокации цепь мРНК передвигается
не на три нуклеотида,
а на два или на четыре. Результаты такой ошибки более серьезны: вся последующая
кодирующая последовательность триплетов мРНК окажется не в фазе, так что синтезируемая
полипептидная цепь не будет иметь ничего общего с нормальным продуктом. Экспрессия генов
во
всех живых организмах находится под контролем разнообразных регуляторных механизмов.
Регуляция генной активности на уровне транскрипции является наиб. изученной.
Менее изучена, хотя и исключительно важна, особенно у высших многоклеточных
организмов, регуляция на уровне Т. Матричная РНК, поступившая в цитоплазму,
может в течение определенного времени вовсе не вовлекаться в синтез белка; разл.
мРНК, вовлеченные в синтез белка, могут транслироваться с очень разной скоростью;
транслируемая мРНК под действием определенных регуляторных факторов может оказаться
нетранслируемой. Ярким примером такого рода
регуляторных переключений являются события, происходящие в ответ на тепловой
шок. Процессы клеточной дифференцировки также сопровождаются включением в Т.
новых мРНК, иногда накопленных в цитоплазме заранее, а также изменением скоростей
Т. и выключением нек-рых мРНК из Т. Регуляция синтеза белков на уровне Т. играет
важную роль у всех организмов, включая бактерии, в координации продукции разл.
белков в клетке и поддержании их правильных стехиометрич. соотношений (это особенно
касается поддержания стехиометрии синтеза субъединиц сложных белков). Регуляция Т. может осуществляться
на всех трех стадиях -инициации, элонгации и терминации. Она м. б. тотальной,
когда ослабление или усиление Т. касается всех мРНК, или избирательной, если
имеет место специфич. подавление или стимуляция использования данной мРНК для
Т. Примером механизма тотального
подавления инициации Т. [у эукариот в ответ на недостаток тема или железа в
ретикулоцитах (вид эритроцитов), на совместное действие интерферона и вирусной
двухцепочечной РНК в ряде др. животных клеток, на увеличение уровня окисленного
глута-тиона и на нек-рые др. неблагоприятные воздействия] является индукция
или активация спец. протеинкиназы, фосфо-рилирующей фактор инициации 2 (сIF-2).
В результате фосфорилирования eIF-2 не инактивируется, но вступает в более тесную
ассоциацию с другим, помогающим ему фактором инициации (eIF-2B), и тем самым
снижает оборачиваемости eIF-2 в процессе инициации. Избирательная регуляция
инициации базируется на трех осн. механизмах: 1) конкуренции разл. мРНК, обладающих
разной "силой" их инициаторных нуклеотидных последовательностей
(как правило, последовательностей, предшествующих инициаторному кодону), за
связывание с малой рибосомной субчастицей и факторами инициации; 2)изме-нениях
пространств. структуры инициаторного района мРНК, от к-рой зависит стерич. доступность
инициаторной последовательности для взаимод. с малой рибосомной субчастицей
и факторами инициации; 3) действии спец. белков (репрессоров Т.), специфически
связывающихся с инициа-торными районами определенных мРНК и делающих их недоступными
для инициации. Скорость Т. в процессе
элонгации также может подвергаться регуляции, как тотальной, так и избирательной.
Макс. скорости чтения мРНК рибосомами в отсутствие всяких ограничивающих факторов
у прокариот и эукариот составляют соотв. 50 и 30 нуклеотидных остатков в секунду
(при 37 °С); регуляторные воздействия могут уменьшить ее до 3-10 нуклеотидов
в секунду. Тотальная регуляция скорости
элонгации у эукариот может осуществляться через фосфорилирование фактора элонгации
2 (EF-2). Показано, что существует спец. Са2+ -калмо-дулинзависимая
протеинкиназа (EF-2-киназа), к-рая может активироваться при разл. физиол. воздействиях
и фосфори-лировать ту или иную часть EF-2 в клетке; фосфорилиро-ванный EF-2
теряет способность эффективно катализировать транслокацию, и элонгация на всех
мРНК замедляется. Фосфатазы или их активация легко обращают этот эффект. 
Тотальная регуляция скорости
элонгации отмечается по ходу прохождения разл. стадий клеточной диффсренциров-ки,
под действием ряда гормонов, при вирусных инфекциях. Лит.: Шапвиль Ф.,
Энни А.-Л., Биосинтез белка, пер. с франц., М., 1977; Спирин А. С., Молекулярная
биология. Структура рибосомы и биосинтез белка, М., 1986. А. С. Спирт
. |
Поделитесь с друзьями:
Вы можете поставить ссылку на это слово:
будет выглядеть так: ТРАНСЛЯЦИЯ
будет выглядеть так: Что такое ТРАНСЛЯЦИЯ